


Plants don’t have a circulatory system like animals, but they do have a sophisticated transport system for carrying water and dissolved solutes to different parts of the plant, often over large distances.

Epidermis : One cell thick. In young plants the epidermis cells may secrete a waterproof cuticle, and in older plants the epidermis may be absent, replaced by bark.
Cortex : Composed of various “packing” cells, to give young plants strength and flexibility, and are the source of plant fibres such as sisal and hemp.
Vascular Tissue : This contains the phloem and xylem tissue, which grow out from the cambium. In dicot plants (the broad-leafed plants), the vascular tissue is arranged in vascular bundles, with phloem on the outside and xylem on the inside. In older plants the xylem bundles fuse together to form the bulk of the stem.
Pith : The central region of a stem, used for food storage in young plants. It may be absent in older plants (i.e. they’re hollow).

Epidermis : A single layer of cells often with long extensions called root hairs, which increase the surface area enormously. A single plant may have 1010 root hairs.
Cortex : A thick layer of packing cells often containing stored starch.
Endodermis : A single layer of tightly-packed cells containing a waterproof layer called the casparian strip. This prevents the movement of water between the cells.
Vascular Tissue : This contains xylem and phloem cells, which are continuous with the stem vascular bundles. The arrangement is different, and the xylem usually forms a star shape with 2-6 arms.
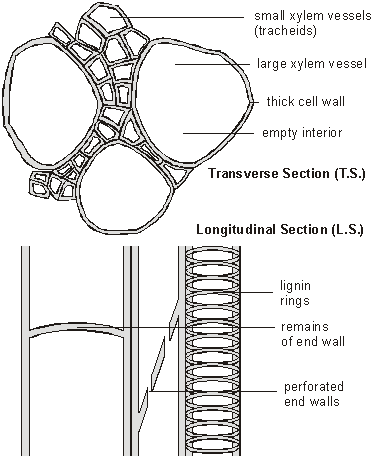
Xylem Tissue
Xylem tissue is composed of dead cells joined together to form long empty tubes. Different kinds of cells form wide and narrow tubes, and the end cells walls are either full of holes, or are absent completely. Before death the cells form thick cell walls containing lignin, which is often laid down in rings or helices, giving these cells a very characteristic appearance under the microscope. Lignin makes the xylem vessels very strong, so that they don’t collapse under pressure, and they also make woody stems strong.

Phloem Tissue
Phloem tissue is composed of sieve tube cells, which form long columns with holes in their end walls called sieve plates. These cells are alive, but they lose their nuclei and other organelles, and their cytoplasm is reduced to strands around the edge of the cells. These cytoplasmic strands pass through the holes in the sieve plates, so forming continuous filaments. The centre of these tubes is empty. Each sieve tube cell is associated with one or more companion cells, normal cells with nuclei and organelles. These companion cells are connected to the sieve tube cells by plasmodesmata, and provide them with proteins, ATP and other nutrients.
Water Transport in Plants
Vast amounts of water pass through plants. A large tree can use water at a rate of 1 dm³ min-1. Only 1% of this water is used by the plant cells for photosynthesis and turgor, and the remaining 99% evaporates from the leaves and is lost to the atmosphere. This evaporation from leaves is called transpiration.
The movement of water through a plant can be split into three sections: through the roots, stem and leaves:

Water moves through the root by two paths:
The Symplast pathway consist of the living cytoplasms of the cells in the root (10%). Water is absorbed into the root hair cells by osmosis, since the cells have a lower water potential that the water in the soil. Water then diffuses from the epidermis through the root to the xylem down a water potential gradient. The cytoplasms of all the cells in the root are connected by plasmodesmata through holes in the cell walls, so there are no further membranes to cross until the water reaches the xylem, and so no further osmosis. The Apoplast pathway consists of the cell walls between cells (90%). The cell walls are quite thick and very open, so water can easily diffuse through cell walls without having to cross any cell membranes by osmosis. However the apoplast pathway stops at the endodermis because of the waterproof casparian strip, which seals the cell walls. At this point water has to cross the cell membrane by osmosis and enter the symplast. This allows the plant to have some control over the uptake of water into the xylem.
The uptake of water by osmosis actually produces a force that pushes water up the xylem. This force is called root pressure, which can be measured by placing a manometer over a cut stem, and is of the order of 100 kPa (about 1 atmosphere). This helps to push the water a few centimetres up short and young stems, but is nowhere near enough pressure to force water up a long stem or a tree. Root pressure is the cause of guttation, sometimes seen on wet mornings, when drops of water are forced out of the ends of leaves.
Movement through the Stem
The xylem vessels form continuous pipes from the roots to the leaves. Water can move up through these pipes at a rate of 8m h-1, and can reach a height of over 100m. Since the xylem vessels are dead, open tubes, no osmosis can occur within them. The driving force for the movement is transpiration in the leaves. This causes low pressure in the leaves, so water is sucked up the stem to replace the lost water. The column of water in the xylem vessels is therefore under tension (a stretching force). Fortunately water has a high tensile strength due to the tendency of water molecules to stick together by hydrogen bonding (cohesion), so the water column does not break under the tension force. This mechanism of pulling water up a stem is sometimes called the cohesion-tension mechanism.
The very strong lignin walls of the xylem vessels stops them collapsing under the suction pressure, but in fact the xylem vessels (and even whole stems and trunks) do shrink slightly during the day when transpiration is maximum.
Movement through the Leaves

The xylem vessels ramify in the leaves to form a branching system of fine vessels called leaf veins. Water diffuses from the xylem vessels in the veins through the adjacent cells down its water potential gradient. As in the roots, it uses the symplast pathway through the living cytoplasm and the apoplast pathway through the non-living cell walls. Water evaporates from the spongy cells into the sub-stomatal air space, and diffuses out through the stomata.

Factors affecting Transpiration
The rate of transpiration can be measured in the lab using a potometer (“drinking meter”):

A potometer actually measures the rate of water uptake by the cut stem, not the rate of transpiration; and these two are not always the same. During the day plants often transpire more water than they take up (i.e. they lose water and may wilt), and during the night plants may take up more water than they transpire (i.e. they store water and become turgid). The difference can be important for a large tree, but for a small shoot in a potometer the difference is usually trivial and can be ignored.
The potometer can be used to investigate how various environmental factors affect the rate of transpiration.
Light : Light stimulates the stomata to open allowing gas exchange for photosynthesis, and as a side effect this also increases transpiration. This is a problem for some plants as they may lose water during the day and wilt.
Temperature : High temperature increases the rate of evaporation of water from the spongy cells, and reduces air humidity, so transpiration increases.
Humidity : High humidity means a higher water potential in the air, so a lower water potential gradient between the leaf and the air, so less evaporation.
Air movements : Wind blows away saturated air from around stomata, replacing it with drier air, so increasing the water potential gradient and increasing transpiration.
Many plants are able to control their stomata, and if they are losing too much water and their cells are wilting, they can close their stomata, reducing transpiration and water loss. So long periods of light, heat, or dry air could result in a decrease in transpiration when the stomata close.








0 comments:
Post a Comment